Mycobacterium tuberculosisESAT-6 antigen immunogenicity in Owl Monkeys
Martha N. Calderon, Carlos Parra-López, Rosalba Alfonso, Paola Barato, Diana Giraldo, Martha L. Pinto, Manuel A. Patarroyo.
Molecular Biology Department, Fundación Instituto de Inmunología de Colombia Universidad Nacional de Colombia
Corresponding author: mapatarr@fidic.org.co
Several ESAT-6-based vaccines have been widely studied in different animal models, presenting potent ability to induce both cellular and humoral responses. As ESAT-6 is a well-characterized mycobacterial antigen, its capacity to induce immune responses in a nonhuman primate model has been evaluated. The immunization of recombinant ESAT-6 (rESAT-6) in Aotus nancymaae monkeys has elicited a strong cellular response, not just to rESAT-6, but also to the native protein present in Mycobacterium tuberculosis culture filtrate proteins measured as [3H]thymidine incorporation in lymphocyte proliferation assays. High humoral response was also observed, having antibody titers of 1:12,800 directed towards rESAT-6. The protein.s multi-epitope nature was further demonstrated since several peptide sequences were specifically recognized at both cellular and humoral level. The high immunogenicity observed, as well as the relatively high characterization of the Aotus. immune system at molecular level, are two advantage to propose Aotus nancymaae as animal model for studying M. tuberculosis infection; however, our results reveal an animal-to-animal variation in response to vaccination, this could be a disadvantage.
Keywords: Owl monkey, animal model, Mycobacterium tuberculosis ESAT-6, immune response.
La única vacuna disponible contra la tuberculosis es la cepa Mycobacterium bovis BCG, que ofrece una eficacia protectiva variable (0%-80%), siendo urgente un nuevo agente profiláctico. Se han evaluado diversos candidatos a vacuna contra este patógeno, en los modelos animales de experimentación convencionales (murino, cobayo, conejo), obteniéndose información básica sobre el efecto de la vacuna en la carga bacterial frente a un reto infeccioso, así como también la reducción o prevención de la patología en los pulmones u otros órganos blanco; además de los aspectos relacionados con la respuesta inmune hacia el Mycobacterium tuberculosis. Los primates no humanos tienen ventajas sobre los modelos convencionales en la evaluación de vacunas, de hecho se ha verificado el comportamiento de agentes terapéuticos en humanos después de haber sido medida la capacidad protectiva de éstos en monos con tuberculosis inducida. Los primates más estudiados en la infección por micobacterias son el cynomulgus, y el rhesus, observándose que estos animales mantienen la infección en un estado subclínico, muy similar a la tuberculosis humana donde el 90% de la población infectada mantiene la infección en un estado latente.
Dado que el modelo animal debe semejar el comportamiento de las proteínas estudiadas en el ser humano, el mono Aotus puede representar ventajas en investigación de tuberculosis por ser un primate con aproximadamente un 90% de similitud al humano en cuanto a las moléculas del sistema inmune estudiadas hasta hoy. La proteína ESAT-6 de (early secretory antigenic target 6 kD) de Mycobacterium tuberculosis es un componente minoritario del filtrado de cultivo de corto tiempo (CFP), ha sido genética y químicamente caracterizada e induce una potente respuesta inmunogénica del tipo TH1. Este antígeno es secretado durante la fase inicial de crecimiento siendo fuertemente reconocido por animales y humanos infectados por Mycobacterium tuberculosis, este hecho hace que su inclusión en una futura vacuna anti-tuberculosis por subunidades y en pruebas de inmunodiagnóstico.
En este trabajo se estudió la respuesta inmune del mono Aotus frente al antígeno micobacteriano ESAT-6 en forma recombinante (rESAT-6). El antígeno fue inmunizado junto con el adyuvante Montanide 720 produciendo una fuerte respuesta humoral y celular, no solo hacia rESAT-6 sino también hacia la proteína nativa presente en el filtrado de medio de cultivo de Mycobacterium tuberculosis. La respuesta celular fue medida por la incorporación de [3H]timidina en ensayos de linfoproliferación. La respuesta humoral se analizó por ensayos de ELISA e inmunoblot, obteniéndose altos títulos de anticuerpos (hasta de 1:12,800) dirigidos rESAT- 6. Se demostró la naturaleza multiepitope de este antígeno ya que en general péptidos que mapean la proteína fueron específicamente reconocidos tanto a nivel celular como humoral. Se observó además variación en la respuesta a la vacunación entre animal y animal, siendo una desventaja para un modelo de experimentación en cuanto a la predicción de la respuesta. Este estudio se constituye en un primer paso de evaluación del mono Aotus como modelo animal en tuberculosis.
Palabras claves: Mono Aotus, modelo animal, Mycobacterium tuberculosis ESAT-6, respuesta inmune.
The only available anti-tuberculosis vaccine is based on the Mycobacterium bovis BCG strain giving variable protective efficacy (0%-80%), making developing a new prophylactic agent an urgent priority (1-3). Great advances have been made in knowledge regarding Mycobacterium tuberculosis, including deciphering the genome.s sequence (4), bioinformatics analysis in silico, proteomic studies directed towards characterizing and determining mycobacterial proteinfunction and immunogenicity (5-8), the mycobacterium.s cell wall structure and morphology (9,10) as well as the role of the main cellular populations, cytokines and chemokines involved in Mycobacterium tuberculosis infection (11,12).
Possible vaccine candidates against this pathogen have been developed having diverse characteristics such as recombinant vaccines, auxotroph vaccines and DNA vaccines (13-19). Orme I. et. al have reported that about 50% of the 170 candidates or combinations of candidates which have been tested fall into the “subunit vaccine category”, reflecting increasing knowledge regarding protein make-up of the bacillus and increased access to techniques helping identify immunogenic proteins (20). These candidate vaccines have been especially evaluated in mouse, guinea-pig and rabbit models which have provided basic information about the vaccine.s effect on bacterial load regarding infectious challenge and reducing or preventing pathology in the lungs and other target organs as well as some aspects related to the immune response to Mycobacterium tuberculosis (21-25).
Nonhuman primates appear to have significant advantages over conventional laboratory animals in terms of modelling pulmonary tuberculosis for purposes of vaccine evaluation (26). The usefulness of tuberculosis induced in monkeys for developing different therapeutic agents has been verified when predicting these drugs. behaviour in humans (27,28). Non-human primate studies have demonstrated that these animals are susceptible to tuberculosis when challenged (29,30).
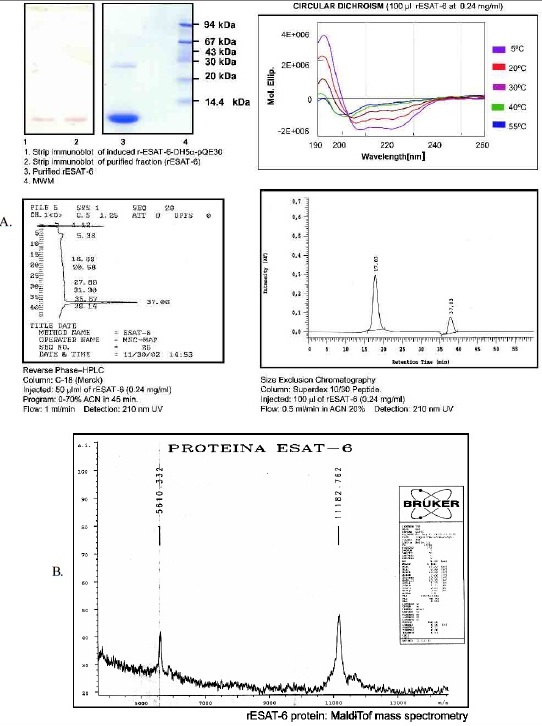
Figure 1. rESAT-6 characterizing. (A) Clonation, expression and purification of rESAT-6: gel electrophoresis, immunoblot, HPLC chromatography and circular dichroism. (B) Maldi-Tof mass spectrometry.
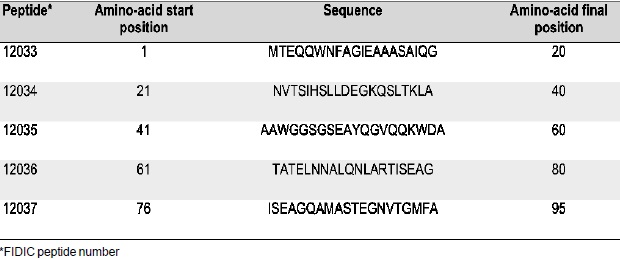
Table I. ESAT-6 synthetic peptide sequences
According to reports, a significant percentage of Macaca fasicularis (cynomolgus monkeys) challenged with low M. tuberculosis doses were able to maintain the infection in a sub-clinical state, very similar to human tuberculosis where 90% of the infected population have latent infection. Other studies have shown that non-human primates can be partially protected by vaccination with BCG and display a delayed-type hypersensitivity (DTH) to prior contacts with mycobacterial antigens (27,31-32). M. tuberculosis culture filtrate protein (CFP) immunogenicity and safety in rhesus and baboon monkeys have been investigated; results showed a short-lived immunity at both humoral and cellular responses, with low IFNg production levels (33,34).
ESAT-6 (early secretory antigenic target 6 kD) is a short-term culture filtrate component; it is a genetic and chemically characterized protein inducing potent TH1 responses (35-41). This antigen is secreted during mycobacterial initial growth phase, being strongly recognized in different species infected with M. tuberculosis (humans, bovines and mice); this means that its inclusion in a future anti-tuberculosis subunit vaccine and in current immuno-diagnostic tests is well-founded (39,42-44).
The Aotus owl monkey.s immune response to rESAT-6 is studied in this work. Immunization with the recombinant antigen together with Montanide 720 adjuvant produces very good humoral and cell-specific responses, even for a long period of time. It was observed at humoral immune response that ESAT-6 protein present in Mycobacterium tuberculosis CFP was recognized by Aotus sera immunized with rESAT-6. Efficient in vitro stimulation of lymphocytes with both recombinant protein and CFP was determined in proliferation assays, indicating that a cell-mediated immune response was being generated.
Immunogen and antigens. rESAT-6 was expressed in DH5a-pQE30 prokaryote system (QIAGEN), was purified by affinity chromatography using a Metal Chelate Affinity-Ni2+ resin and imidazol gradient. rESAT-6 was characterized by immunoblot, HPLC (reverse
phase and size exclusion chromatography), Maldi-Tof mass spectrometry and circular dichroism, Figure 1.
The Mycobacterium bovis BCG live vaccine (Lyophilized, Inter Vax, Canada) was also used as reactivity control for in vivo experiments. Crude Mycobacterium tuberculosis H37Rv culture filtrate protein (CFP) produced by the National Institutes of Health, National Institute of Allergy and Infectious Diseases (Contract N01-AI-75320, kindly donated by Dr. J. Belisle at Colorado State University) was used in the in vitro immunological assays.
Table II. Immunized Aotus monkey groups

Immunogens were dissolved in PBS (I, II, and III) and emulsified 30:70 with Montanide ISA720 adjuvant (I and III). M. bovis BCG was not emulsified in Montanide ISA720 adjuvant
6 protein 20-amino-acid long peptides were synthesized by solid phase methodology (53), purified by HPLC and characterized by mass spectrometry. Peptide numbers and sequences are shown in Table I.
Animals.
A total of 15 healthy Aotus nancymaae owl monkeys (having been previously orally deparasitized with Ivermectin) were selected based on immunoblots of the sera analyzed having minimum or null mycobacterial recognition (Mycobacterium tuberculosis H37Rv lysate). Monkeys were kept at our institute.s experimental primate station in Leticia (Amazon) in accordance with CCAC (Committee on Care and Use of Laboratory Animals, US) guidelines and Colombian National Institute of Health guidelines for the use of laboratory animals, supervised by the ColombianWildlife Corporation (CorpoAmazonia).
The presence of mycobacteria in monkeys housed at the colony was evaluated by randomly taken samples. One hundred samples from different sources, gastric lavage, faecal material, soil, food and drinkingwater were thus analyzed. Advantage was taken of the anaesthesia (35mg/kg weight ketamin, 0.1mg/Kg atropine and 1 mg/Kg diazepam, applied intramuscularly) required for some procedures carried out on the 30 Aotus for also performing gastric lavage using a size 6 Levin probe (54). The samples were decontaminated by Petroff.s technique (55). Ziehl. Neelsen (ZN) staining, and culturing in Bactec MIGT960 system and solid medium (Middlebrook 7H10) (for 6 weeks at 37°C) were then carried out.
The tuberculin test was done in the eyelids of 15 Aotus previously positive for acid-fast bacilli (AFB) and/or rejected from the study during the selection process due to having some type of specific mycobacterial antigen recognition. A 100ml tuberculin dose (PPD RT23 SSI, Swtatens Serum Institute, Copenhagen) was intra-dermally administered.
Immunization.
Five monkeys were selected for each immunogen (Table II). 100mg rESAT-6 in PBS (137 mM NaCl, 2.7 mM KCl, 4.3 mM Na2HPO4 and 1.4 mM KH2P04) per monkey-dose were applied to the first group. The second group received only Mycobacterium bovis BCG; this was reconstituted in PBS, applied at 50mg per dose. The third group corresponded to the placebo (PBS plus adjuvant). Immunogens were emulsified with Montanide ISA 720 adjuvant (SEPPIC-France) in a 70:30 ratio by sonication when being applied.
Aotus were immunized subcutaneously with 200ml emulsion in the abdomen three times at 30 day intervals (days 0, 30 and 60). Blood was collected immediately prior to the first injection, on day 30 and every month on three occasions. Monkeys were physically monitored for 180 days for developing local and systemic reactions.
Antibody response.
For enzyme-linked Immunosorbent assays (ELISAs), the antigens were diluted in PBS at 10mg/ well (CFP) and 1 mg/well (rESAT-6 and peptides) and used for coating microtiter wells; the wells were blocked with 3% BSA. Serum samples (previously adsorbed with E. coli lysate proteins) were added at diferent dilutions; horseradish peroxidase (HRP)- conjugated goat anti-Aotus IgG (produced by FIDIC) was added to each well as second antibody. The reaction was developed using TMB Microwell Peroxidase Substrate System, A620nm was read on a Labsystems Multiskan MJ ELISA reader (56).
For Western Blot analysis, CFP were separated by sodium dodecylsulphate-polyacrylamide gel electrophoresis (SDS-PAGE), under reducing conditions on a 10-15% gradient gel. rESAT-6 was separated by 14% SDS-PAGE in Tris-Tricine system. The gels were electrophoretically transferred to nitrocellulose membrane (Trans-Blot Bio Rad).
Blocking buffer (1% nonfat milk-TBS (10mM TrisHCl pH7.5, 150mM NaCl)-0.05% Tween 20) was used for saturating non-specific sites. Single nitrocellulose strips with nearly 50mg CFP or 1 mg rESAT-6 were incubated with monkey serum diluted 1:100 in blocking buffer. Strips were incubated with Alkaline phosphatase (AP)-labelled goat anti-Aotus IgG, BCIP/ NBT (Promega) was used for detection (57).
Cellular proliferative response.
Monkeys were splenectomized on day 150 after the study was begun. Lymphocytes were obtained by preparing single-cell suspensions from spleens and then purified by Ficoll-Hypaque (ICN Lympho separation medium) gradient centrifugation. Cells were washed and suspended in RPMI 1640 medium containing HEPES, L-glutamine, antibiotic-antimycotic and 20% autologous serum. 100ml cell suspension containing 106 cells were plated onto 96-well flatbottom plates, together with antigen (13). CFP (10mg/ ml), rESAT-6 or peptides (1mg/ml) were used as antigens and 5mg/ml phytohemagglutinin (PHA, DIFCO) or RPMI 1640 as positive control and nonstimulated control respectively. Each stimulation was performed in triplicate wells. Plates were incubated at 37°C in 5% CO2 for 72 h. [3H]thymidine (1mCi/well; Amersham) was added for the final 16-18 hours incubation. Cells were harvested onto filters (Glass Fibre Strips 240-1, Cambridge Tech.) by using a cell harvester; filters were dried and cells were counted in a scintillation counter. Results are reported as stimulation index (SI): a two-fold increase in counts per minute over non-stimulated control were considered positive .
Results
Mycobacterial presence.
No signs of tuberculous infection were observed; few alcohol-resistant bacilli were found in those randomly-chosen samples taken; only 8 positive specimens were isolated in the population analyzed in BACTEC culture and verified by ZN: 4 came from
gastric lavage, 1 from soil and 3 from faecal material. DNA isolated from cultured samples was further analyzed by PCR (54), being negative for M. tuberculosis complex mycobacteria (data not shown). The tuberculin test was done in the eyelids of 15 Aotus
previously positive for AFB and/or rejected from the study during the selection process due to having some type of specific mycobacterial antigen recognition; no induration or positive reaction was detected 24 to 72 hours following application.
Aotus selection and immunization.
Two parameters were taken into account in the process of selecting animals for this study: optimum state of health (indicated by clinical evaluation) and minimum or null sera recognition of M. tuberculosis H37Rv proteins inWestern blot assay. Sera adsorption with E. coli proteins allowed specific recognition of mycobacterial proteins which is why this procedure was done prior to the immunoblot and ELISA analyses.
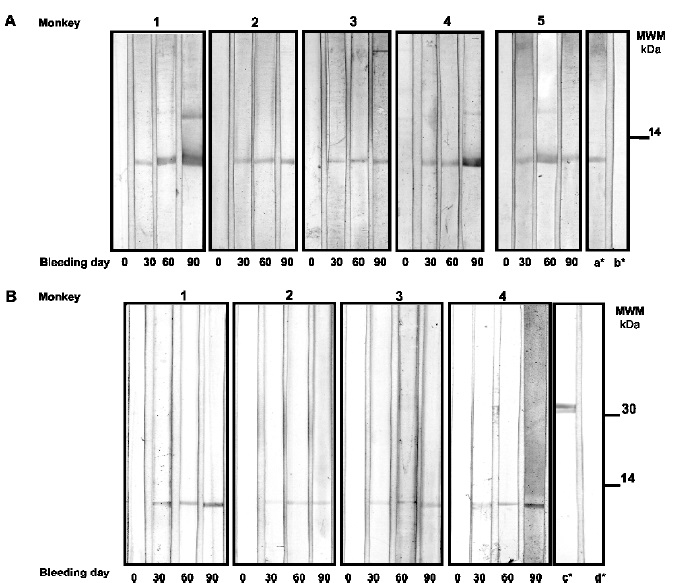
Figure 2. Western blot analysis showing the binding of antibodies from immunized Aotus monkeys with rESAT-6. (A) Results of immunodetection in rESAT-6. (B) Mycobacterium tuberculosis crude CFP (B). Primary antibodies 1:100 dilution sera. Assay controls: a* strip incubated with anti-polyHistidine (Sigma), then anti-mouse IgG-PA (ICN.); c* strip incubated with anti-Ag85B, then anti-rabbit IgG-PA (ICN); b* and c* reagent control for A and B, respectively
Monkeys were immunized on three times, presenting satisfactory clinical analysis. No evidence was seen of nodules or erythema at the application site in animals immunized with rESAT-6; similar results were obtained in the placebo monkeys. Aotus group II received a single dose of BCG; nodulations were observed which were immobile to the touch at the application site compromising skin and subcutaneous tissue, Table II.
Humoral immune response.
Immunoblot and ELISA were used for determining rESAT-6 and CPF antibody recognition by monkey sera. Antisera obtained from group I animals at various times after the first and subsequent immunizations were able to strongly bind rESAT-6 dose-dependently for most animals, Figure 2A. Except for Monkey 5, which displayed very low recognition (data not shown), native ESAT-6 present in CFP was also recognized, even from the first bleeding. There was less recognition intensity, however, out of the whole protein pattern found in CFP (more than 100 proteins), specificity was confirmed as the only band being recognized had the same molecular mass as ESAT-6, Figure 2B.
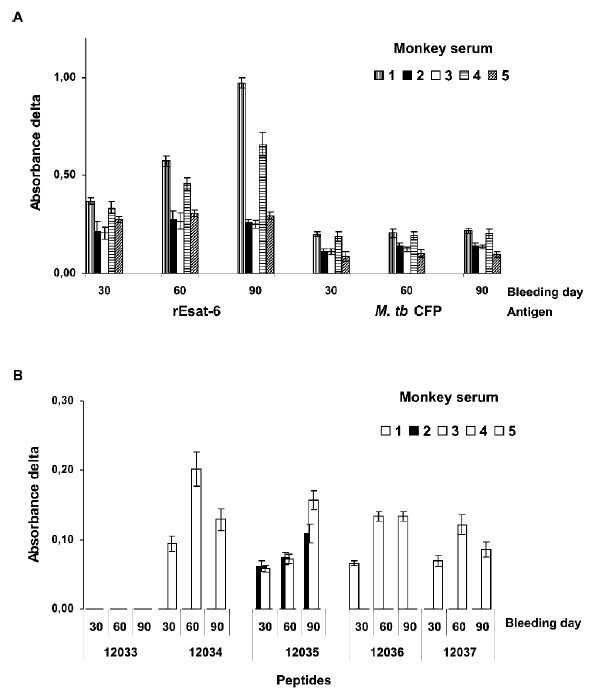
Figure 3. ELISA assay for IgG antibodies generated by immunization with rESAT. (A) CFP and rESAT. (B) 5 peptides mapping ESAT-6. 620nm absorbance delta, corresponding to the value obtained in each bleeding less pre-immune sera absorbance of each monkey regarding each antigen. Primary antibodies 1:100 dilution. Each bar represents the mean of triplicate values.
ELISA assays corroborated the foregoing; it was determined that the rESAT-6 antigen binding titer became progressively increased until obtaining titers between 1:800 and 1:12,800 for bleeding III (after 90 days). Antibody titers measured in CFP were generally constant in the different immunizations, 1:100 being the lowest value (Figure 3A). Studying the different peptide sequences mapping the protein showed varied recognition for the different animals.
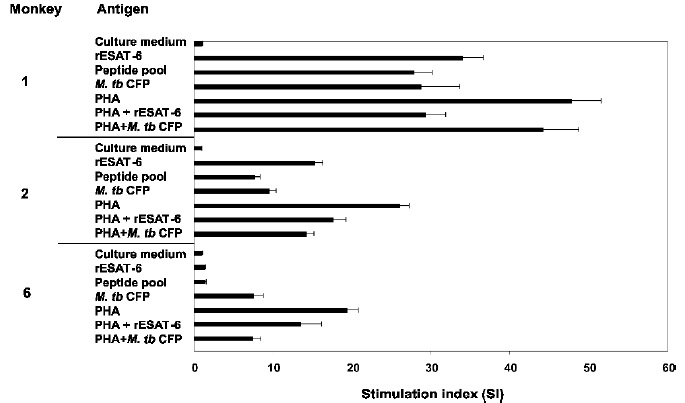
Figure 4. Proliferative responses of spleen-derived lymphocytes from Aotus immunized with rESAT-6 (Monkeys 1 and 2) and BCG (Monkey 6). [3H]thymidine incorporation in response to in vitro stimulation with rESAT-6, the pool of ESAT-6 peptides, CFP and PHA (positive control) were measured 90 days after final immunization. Data are reported as SIs (fold stimulation over that of cells incubated in just medium). Results shown are representative of the immunized population. Each bar represents the mean of triplicate values ± sem
Except for Monkey 5 (having no appreciable antibody titer values to any particular peptide), the other antisera recognized sequences throughout the whole protein, amino-terminal portion (NH2-ter) not being recognized (Figure 3B). No significant rESAT-6 antibody titers were observed when sera from animals immunized with BCG or placebo were tested (data not shown).
Cellular immune response.
Cellular proliferative responses were assessed using animals. spleen lymphocytes. Figure 4 shows proliferation levels induced by rESAT-6, the pool of peptides mapping the protein, CFP and PHA in two representative experiments of rESAT-6 immunized animals (Monkeys 1 and 2).All immunized monkeys developed a proliferative cellular response to CFP, including monkeys immunized with BCG; Figure 4 shows Monkey 6 lymphoproliferation results which were typical of Aotus immunized withBCG. In all cases proliferation induced by PHA was greater than that found with the other antigens (recombinant and peptides), including that displayed by placebo monkey lymphocytes. Recombinant protein and peptide pool stimulation was specific in those animals immunized with just rESAT-6. The results show strong stimulation of antigens, even though being different for each monkey. SI values ranged from 15 to 34 for rESAT-6; 8 to 28 for the peptide pool; and 10 to 29 for CFP.
The proliferative response of cells from rESAT-6 immunized animals stimulated with peptides mapping the protein was lower than that observed when stimulation with the recombinant protein was carried out, Table IV. Contrary to what happens with humoral response, several monkeys recognized the protein.s NH2-ter portion (peptide 12033) simulta-neously with the carboxy-terminal region (COOH-ter), as observed in Monkeys 2 and 3. The other Aotus recognized the (COOH-ter) region with peptides 12036 and 12037. None of the peptides tested was recognized by Monkey 5. Cells stimulated with peptides from the protein.s middle region (12034 and 12035) did not present any appreciable proliferation.
Table III. rESAT-6 binding antibody titers obtained by ELISA assay
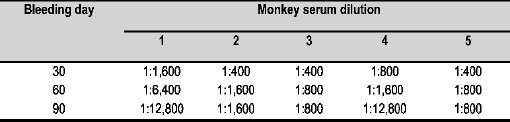
Table IV. ESAT-6 peptide-stimulated splenocyte proliferation assays
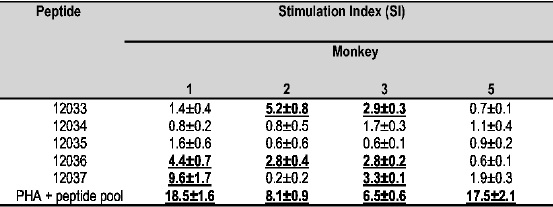
Monkey 4 died before the experiment ended. Positive values were considered as having SI>2 (shown in bold and underlined). Each result represents the mean of triplicate values ± sem.
Immunogenicity and protection-inducing levels have been explored in diverse preparations such as DNA and recombinant ESAT-6 vaccines (13-14,17,64-65). These candidates have been able to induce cellular TH1 response and protection similar to that presented by BCG. Subunit vaccines (in which ESAT-6 potential has been evaluated) have also been reported; determining the modulating effect of the immune response of the adjuvant used in the immunization process, depending on the antigen. One study has shown that the ESAT-6-DDA combination (dimethyl dioctadecyl ammoniumbromide) does not induce any appreciable immune response in the murine model. When MPL (monophosphoryl lipid A) is incorporated into the preparation in different combinations, strong adjuvant activity can be observed for both cellular and humoral immune responses (42). The data presented in different reports indicate that protective immunity determined for different ESAT-6 vaccines is at the same level as BCG, a relevant result for a single M. tuberculosis antigen; these results show high humoral and cellular response when recombinant protein was administered using Montanide 720 as adjuvant. Montanide 720 is a metabolizable mannide oleate in oil solution adjuvant which has been recently approved for use in humans. Aotus immunized with rESAT-6 or the placebo did not present nodules (as observed with animals immunized with BCG).
Different reports have indicated that ESAT-6 contains numerous epitopes recognized by a high percentage of individuals, depending on the sensitivity of the assay used (59,61,66). Different peptide sequences involved in humoral recognition were observed in Aotus monkeys, where almost the whole protein participates in antibody production. Interestingly none of the immunized animals. Sera recognized the recombinant antigen NH2-ter portion. Likewise, low recognition of ESAT-6 NH2-ter region peptides by antibodies in tuberculous non-human primate sera (cynomulgus macaques, rhesus macaques, African Green monkeys) has been recently demonstrated in a study using overlapping peptides spanning the ESAT-6 protein sequence, while an important recognition towards the COOH-ter portion is observed even early in infection (63). At cellular response level, sequences mainly comprising the COOHter fraction have induced proliferation in all immunized monkeys, while two of the immunized monkeys are able to respond towards the NH2-ter also.
Similar results have been reported in previous studies using human cells (41,66-68). Cellular and humoral in vitro immune response stimulation were lower whenever peptides mapping ESAT-6 protein were used, suggesting that either conformational epitopes were present or that some epitopes were missing due to the use of non-overlapped peptides.
The Aotus nancymaae monkey has been recommended by the World Health Organization (WHO) as a model for malaria vaccine candidate evaluation (69-70). This has favored immune system molecule research leading to a high percentage of homology with humans being elucidated. Recent data has shown 78% to 95% homology for MHC-II molecules (histocompatibility Class-II complex) (45- 52,71) which have been declared as having great importance in tuberculosis infection (23-24). Lymphocytes. proliferative in vitro response to mitogens in Aotus nancymaae has been analyzed, showing that it varies from monkey to monkey depending on the particular mitogen assessed (72); this was also seen in the present work. Varied cellular and humoral recognition was determined amongst different monkeys and even to different antigens; however, the results show that positive recognition values are antigen-specific, having no ambiguity regarding immune response.
It has been reported that all primates are susceptible to tuberculosis, although differences between species exist (e.g. OldWorld primate species such as rhesus monkeys are very sensitive to infection whilst New World species such as squirrel monkeys appear to be less susceptible) (27). There have been few reports recently regarding tuberculosis in NewWorld Monkeys, even more so in the wild. Astudy isolating and identifying mycobacteria in NewWorld primates maintained in captivity identified M. chelonae subsp. abscessus in one of the two Aotus sp. analyzed (54). Few monkeys were found to be positive for mycobacteria in the current study; all isolated mycobacteria were non-pathogenic, although Amazon province (Colombia) is tuberculosis pulmonary disease endemic area (73).
The tuberculin test was done in Aotus previously positive for AFB and/or having some type of mycobacterial antigen recognition; no induration or positive reaction was detected. This result (together with positive recognition of mycobacterial antigens) suggests that such animals were not infected by pathogenic mycobacteria, that low AFB detection in the analyzed population was due to Aotus having been in contact with environmental mycobacteria and that shared epitope cross-recognition was being generated.
Aotus develop a strong immune response when they were immunized with a mycobacterial antigen in a recombinant protein form (rESAT-6) emulsified with Montanide 720 as adjuvant. Immune response had multi-epitope characteristics and was seen to be different among the monkeys analyzed as similar human studies, but this behavior is a disadvantage for a reliable experimental model because de responses are not enough predictable. A significant number animals were analized and not any monkey were found to be positive for pathogenic mycobacteria or positive reaction to tuberculin test.
2. Fine PE: Variation in protection by BCG: implications of and for heterologous immunity. Lancet 346:1339 1345, 1995
3. Ginsberg AM: What.s new in tuberculosis vaccines? Bull World Health Organ 80:483-488, 2002
4 . Cole ST, Brosch R, Parkhill J, Garnier T, Churcher C, Harris D, Gordon SV, Eiglmeier K, Gas S, Barry CE, 3rd, Tekaia F, Badcock K, Basham D, Brown D, Chillingworth T, Connor R, Davies R, Devlin K, Feltwell T, Gentles S, Hamlin N, Holroyd S, Hornsby T, Jagels K, Barrell BG, et al.: Deciphering the biology of Mycobacterium tuberculosis from the complete genome sequence. Nature 393:537-544, 1998
5 . Tekaia F, Gordon SV, Garnier T, Brosch R, Barrell BG, Cole ST: Analysis of the proteome of Mycobacterium tuberculosis in silico. Tuber Lung Dis 79:329-342, 1999
6 . Jungblut PR, Schaible UE, Mollenkopf HJ, Zimny-Arndt U, Raupach B, Mattow J, Halada P, Lamer S, Hagens K, Kaufmann SH: Comparative proteome analysis of Mycobacterium tuberculosis and Mycobacterium bovis BCG strains: towards functional genomics of microbial pathogens. Mol Microbiol 33:1103-1117, 1999
7 . Domenech P, Barry CE, 3rd, Cole ST: Mycobacterium tuberculosis in the post-genomic age. Curr Opin Microbiol 4:28-34, 2001
8. Covert BA, Spencer JS, Orme IM, Belisle JT: The application of proteomics in defining the T cell antigens of Mycobacterium tuberculosis. Proteomics 1:574-586, 2001
9 Brennan PJ: Structure, function, and biogenesis of the cell wall of Mycobacterium tuberculosis. Tuberculosis (Edinb) 83:91-97, 2003
10. Brennan PJ, Besra GS: Structure, function and biogenesis of the mycobacterial cell wall. Biochem Soc Trans 25:188-194, 1997
11. Orme IM, Cooper AM: Cytokine/chemokine cascades in immunity to tuberculosis. Immunol Today 20:307-312, 1999
12. Flynn JL, Chan J: Immunology of tuberculosis. Annu Rev Immunol 19:93-129, 2001
13. Pym AS, Brodin P, Majlessi L, Brosch R, Demangel C, Williams A, Griffiths KE, Marchal G, Leclerc C, Cole ST: Recombinant BCG exporting ESAT-6 confers enhanced protection against tuberculosis. Nat Med 9:533-539, 2003
14. Bao L, Chen W, Zhang H, Wang X: Virulence, immunogenicity, and protective efficacy of two recombinant Mycobacterium bovis bacillus Calmette-Guerin strains expressing the antigen ESAT-6 from Mycobacterium tuberculosis. Infect Immun 71:1656-1661, 2003
15. Jackson M, Phalen SW, Lagranderie M, Ensergueix D, Chavarot P, Marchal G, McMurray DN, Gicquel B, Guilhot C: Persistence and protective efficacy of a Mycobacterium tuberculosis auxotroph vaccine. Infect Immun 67:2867-2873, 1999
16. Kamath AT, Feng CG, Macdonald M, Briscoe H, Britton WJ: Differential protective efficacy of DNA vaccines expressing secreted proteins of Mycobacterium tuberculosis. Infect Immun 67:1702-1707, 1999
17. Li Z, Howard A, Kelley C, Delogu G, Collins F, Morris S: Immunogenicity of DNA vaccines expressing tuberculosis proteins fused to tissue plasminogen activator signal sequences. Infect Immun 67:4780-4786, 1999
18. Huygen K: On the use of DNA vaccines for the prophylaxis of mycobacterial diseases. Infect Immun 71:1613-1621, 2003
19. Nor NM, Musa M: Approaches towards the development of a vaccine against tuberculosis: recombinant BCG and DNA vaccine. Tuberculosis (Edinb) 84:102-109, 2004
20. Orme IM, McMurray DN, Belisle JT: Tuberculosis vaccine development: recent progress. Trends Microbiol 9:115-118, 2001
21. Dannenberg AM Jr, Collins FM: Progressive pulmonary tuberculosis is not due to increasing numbers of viable bacilli in rabbits, mice and guinea pigs, but is due to a continuous host response to mycobacterial products. Tuberculosis (Edinb);81(3):229-42, 2001
22. Baldwin SL, D.Souza C, Roberts AD, Kelly BP, Frank AA, Lui MA, Ulmer JB, Huygen K, McMurray DM, Orme IM: Evaluation of new vaccines in the mouse and guinea pig model of tuberculosis. Infect Immun 66:2951-2959, 1998
23. Scanga CA, Mohan VP, Yu K, Joseph H, Tanaka K, Chan J, Flynn JL: Depletion of CD4(+) T cells causes reactivation of murine persistent tuberculosis despite continued expression of interferon gamma and nitric oxide synthase 2. J Exp Med 192:347-358, 2000
24. Mogues T, Goodrich ME, Ryan L, LaCourse R, North RJ: The relative importance of T cell subsets in immunity and immunopathology of airborne Mycobacterium tuberculosis infection in mice. J Exp Med 193:271-280, 2001
25. McMurray DN: Recent progress in the development and testing of vaccines against human tuberculosis. Int J Parasitol 33:547-554, 2003
26. McMurray DN: A nonhuman primate model for preclinical testing of new tuberculosis vaccines. Clin Infect Dis 30 Suppl 3:S210-212, 2000
27. Good RC: Diseases in Nonhuman Primates. In The Mycobacteria. A Sourcebook. Kubica G, Wayne L (eds). New York, Marcel Dekker. Inc., 1984, pp 903-924
28. VandeBerg JL, Williams-Blangero S: Advantages and limitations of nonhuman primates as animal models in genetic research on complex diseases. J Med Primatol 26:113-119, 1997
29. Walsh GP, Tan EV, dela Cruz EC, Abalos RM, Villahermosa LG, Young LJ, Cellona RV, Nazareno JB, Horwitz MA: The Philippine cynomolgus monkey (Macaca fasicularis) provides a new nonhuman primate model of tuberculosis that resembles human disease. Nat Med 2:430-436, 1996
30. Capuano SV, III, Croix DA, Pawar S, Zinovik A, Myers A, Lin PL, Bissel S, Fuhrman C, Klein E, Flynn JL: Experimental Mycobacterium tuberculosis infection of cynomolgus macaques closely resembles the various manifestations of human M. tuberculosis infection. Infect Immun 71:5831-5844, 2003
31. Langermans JA, Andersen P, van Soolingen D, Vervenne RA, Frost PA, van der Laan T, van Pinxteren LA, van den Hombergh J, Kroon S, Peekel I, Florquin S, Thomas AW: Divergent effect of bacillus Calmette Guerin (BCG) vaccination on Mycobacterium tuberculosis infection in highly related macaque species: implications for primate models in tuberculosis vaccine research. Proc Natl Acad Sci U S A 98:11497 11502, 2001
32. Baer JF: Husbandry and medical management of the owl monkey. In Aotus: The owl monkey Baer JF, Weller RE, Kakoma I (eds). New York, Academic Press Inc., 1994, pp 133-164
33. Attanasio R, Pehler K, McClure HM: Immunogenicity and safety of Mycobacterium tuberculosis culture filtrate proteins in nonhuman primates. Clin Exp Immunol 119:84-91, 2000
34. Pehler K, Brasky KM, Butler TM, Attanasio R: Mycobacterium tuberculosis-secreted protein antigens: immunogenicity in baboons. J Clin Immunol 20:306-316, 2000
35. Berthet FX, Rasmussen PB, Rosenkrands I, Andersen P, Gicquel B: A Mycobacterium tuberculosis operon encoding ESAT-6 and a novel low-molecular-mass culture filtrate protein (CFP-10). Microbiology 144 ( Pt 11):3195-3203, 1998
36. Gey Van Pittius NC, Gamieldien J, Hide W, Brown GD, Siezen RJ, Beyers AD: The ESAT-6 gene cluster of Mycobacterium tuberculosis and other high G+C Gram-positive bacteria. Genome Biol 2:RESEARCH0044, 2001
37. Sorensen AL, Nagai S, Houen G, Andersen P, Andersen AB: Purification and characterization of a low molecular-mass T-cell antigen secreted by Mycobacterium tuberculosis. Infect Immun 63:1710-1717, 1995
38. Harboe M, Oettinger T, Wiker HG, Rosenkrands I, Andersen P: Evidence for occurrence of the ESAT-6 protein in Mycobacterium tuberculosis and virulent Mycobacterium bovis and for its absence in Mycobacterium bovis BCG. Infect Immun 64:16-22, 1996
39. Renshaw PS, Panagiotidou P, Whelan A, Gordon SV, Hewinson RG, Williamson RA, Carr MD: Conclusive evidence that the major Tcell antigens of the Mycobacterium tuberculosis complex ESAT-6 and CFP-10 form a tight, 1:1 complex and characterization of the structural properties of ESAT-6, CFP-10, and the ESAT-6*CFP-10 complex. Implications for pathogenesis and virulence. J Biol Chem 277:21598-21603, 2002
40. Skjot RL, Oettinger T, Rosenkrands I, Ravn P, Brock I, Jacobsen S, Andersen P: Comparative evaluation of low-molecular-mass proteins from Mycobacterium tuberculosis identifies members of the ESAT-6 family as immunodominant T-cell antigens. Infect Immun 68:214-220, 2000
41. Mustafa AS, Oftung F, Amoudy HA, Madi NM, Abal AT, Shaban F, Rosen Krands I, Andersen P: Multiple epitopes from the Mycobacterium tuberculosis ESAT-6 antigen are recognized by antigen-specific human T cell lines. Clin Infect Dis 30 Suppl 3:S201-205, 2000
42. Brandt L, Elhay M, Rosenkrands I, Lindblad EB, Andersen P: ESAT-6 subunit vaccination against Mycobacterium tuberculosis. Infect Immun 68:791-795, 2000
43. Olsen AW, Hansen PR, Holm A, Andersen P: Efficient protection against Mycobacterium tuberculosis by vaccination with a single subdominant epitope from the ESAT-6 antigen. Eur J Immunol 30:1724-1732, 2000
44. Mori T, Sakatani M, Yamagishi F, Takashima T, Kawabe Y, Nagao K, Shigeto E, Harada N, Mitarai S, Okada M, Suzuki K, Inoue Y, Tsuyuguchi K, Sasaki Y, Mazurek GH, Tsuyuguchi I: Specific Detection of Tuberculosis Infection with an Interferongamma Based Assay Using New Antigens. Am J Respir Crit Care Med, 2004
45. Goodman M, Porter CA, Czelusniak J, Page SL, Schneider H, Shoshani J, Gunnell G, Groves CP: Toward a phylogenetic classification of Primates based on DNA evidence complemented by fossil evidence. Mol Phylogenet Evol 9:585-598, 1998
46. Nino-Vasquez JJ, Vogel D, Rodriguez R, Moreno A, Patarroyo ME, Pluschke G, Daubenberger CA: Sequence and diversity of DRB genes of Aotus nancymaae, a primate model for human malaria parasites. Immunogenetics 51:219-230, 2000
47. Daubenberger CA, Salomon M, Vecino W, Hubner B, Troll H, Rodriques R, Patarroyo ME, Pluschke G: Functional and structural similarity of V gamma 9V delta 2 T cells in humans and Aotus monkeys, a primate infection model for Plasmodium falciparum malaria. J Immunol 167:6421-6430, 2001
48. Montoya GE, Vernot JP, Patarroyo ME: Partial characterization of the CD45 phosphatase cDNA in the owl monkey (Aotus vociferans). Am J Primatol 57:1-11, 2002
49. Guerrero JE, Pacheco DP, Suarez CF, Martinez P, Aristizabal F, Moncada CA, Patarroyo ME, Patarroyo MA: Characterizing Tcell receptor gamma-variable gene in Aotus nancymaae owl monkey peripheral blood. Tissue Antigens 62:472-482, 2003
50. Webster D, Hill AV: Progress with new malaria vaccines. Bull World Health Organ 81:902-909, 2003
51. Espejo F, Bermudez A, Torres E, Urquiza M, Rodriguez R, Lopez Y, Patarroyo ME: Shortening and modifying the 1513 MSP-1 peptide.s alpha-helical region induces protection against malaria. Biochem Biophys Res Commun 315:418-427, 2004
52. Rodriguez R, Moreno A, Guzman F, Calvo M, Patarroyo ME: Studies in owl monkeys leading to the development of a synthetic vaccine against the asexual blood stages of Plasmodium falciparum. Am J Trop Med Hyg 43:339-354, 1990
53. Houghten RA: General method for the rapid solid-phase synthesis of large numbers of peptides: specificity of antigenantibody interaction at the level of individual amino acids. Proc Natl Acad Sci U S A 82:5131 5135, 1985
54 Alfonso R, Romero RE, Diaz A, Calderon MN, Urdaneta G, Arce J, Patarroyo ME, Patarroyo MA: Isolation and identification of mycobacteria in New World primates maintained in captivity. Vet Microbiol 98:285-295, 2004
55. Palomino JC, Portaels F: Effects of decontamination methods and culture conditions on viability of Mycobacterium ulcerans in the BACTEC system. J Clin Microbiol 36:402-408, 1998
56. Espinosa AM, Sierra AY, Barrero CA, Cepeda LA, Cantor EM, Lombo TB, Guzman F, Avila SJ, Patarroyo MA: Expression, polymorphism analysis, reticulocyte binding and serological reactivity of two Plasmodium vivax MSP-1 protein recombinant fragments. Vaccine 21:1033-1043, 2003
57. Bollag DM, Edelstein SJ: Immunoblotting. In Protein Methods. (ed). New York, John Wiley and sons, Inc, 1993, pp 181-211
58. Andersen P, Andersen AB, Sorensen AL, Nagai S: Recall of longlived immunity to Mycobacterium tuberculosis infection in mice. J Immunol 154:3359-3372, 1995
59. Ulrichs T, Munk ME, Mollenkopf H, Behr-Perst S, Colangeli R, Gennaro ML, Kaufmann SH: Differential T cell responses to Mycobacterium tuberculosis ESAT6 in tuberculosis patients and healthy donors. Eur J Immunol 28:3949-3958, 1998
60. Brock I, Weldingh K, Lillebaek T, Follmann F, Andersen P: Comparison of a New Specific Blood Test and the Skin Test in Tuberculosis Contacts. Am J Respir Crit Care Med, 2004
61. Brandt L, Oettinger T, Holm A, Andersen AB, Andersen P: Key epitopes on the ESAT-6 antigen recognized in mice during the recall of protective immunity to Mycobacterium tuberculosis. J Immunol 157:3527-3533, 1996
62. Pollock JM, Andersen P: Predominant recognition of the ESAT-6 protein in the first phase of interferon with Mycobacterium bovis in cattle. Infect Immun 65:2587-2592, 1997
63. Kanaujia GV, Motzel S, Garcia MA, Andersen P, Gennaro ML: Recognition of ESAT-6 sequences by antibodies in sera of tuberculous nonhuman primates. Clin Diagn Lab Immunol 11:222-226, 2004
64 Morris S, Kelley C, Howard A, Li Z, Collins F: The immunogenicity of single and combination DNA vaccines against tuberculosis. Vaccine 18(20):2155-63, 2000
65. Minion FC, Menon SA, Mahairas GG, Wannemuehler MJ: Enhanced murine antigen-specific gamma interferon and immunoglobulin G2a responses by using mycobacterial ESAT-6 sequences in DNA vaccines. Infect Immun 71:2239-2243, 2003
66. Ravn P, Demissie A, Eguale T, Wondwosson H, Lein D, Amoudy HA, Mustafa AS, Jensen AK, Holm A, Rosenkrands I, Oftung F, Olobo J, von Reyn F, Andersen P: Human T cell responses to the ESAT-6 antigen from Mycobacterium tuberculosis. J Infect Dis 179:637-645, 1999
67. Vincenti D, Carrara S, De Mori P, Pucillo LP, Petrosillo N, Palmieri F, Armignacco O, Ippolito G, Girardi E, Amicosante M, Goletti D: Identification of early secretory antigen target-6 epitopes for the immunodiagnosis of active tuberculosis. Mol Med 9:105-111, 2003
68. Mustafa AS, Shaban FA, Al-Attiyah R, Abal AT, El-Shamy AM, Andersen P, Oftung F: Human Th1 cell lines recognize the Mycobacterium tuberculosis ESAT-6 antigen and its peptides in association with frequently expressed HLA class II molecules. Scand J Immunol 57:125-134, 2003
69. Patarroyo ME, Romero P, Torres ML, Clavijo P, Moreno A, Martinez A, Rodriguez R, Guzman F, Cabezas E: Induction of protective immunity against experimental infection with malaria using synthetic peptides. Nature 328:629-632, 1987
70. Collins WE, Galland GG, Sullivan JS, Morris CL: Selection of different strains of Plasmodium falciparum for testing bloodstage vaccines in Aotus nancymai monkeys. Am J Trop Med Hyg 51:224-232, 1994
71. Diaz D, Naegeli M, Rodriguez R, Nino-Vasquez JJ, Moreno A, Patarroyo ME, Pluschke G, Daubenberger CA: Sequence and diversity of MHC DQA and DQB genes of the owl monkey Aotus nancymaae. Immunogenetics 51:528-537, 2000
72. Pinzon-Charry A, Vernot JP, Rodriguez R, Patarroyo ME: Proliferative response of peripheral blood lymphocytes to mitogens in the owl monkey Aotus nancymae. J Med Primatol 32:31-38, 2003
73. Chaparro PE, García I, Guerrero MI, León CI: Situación de la tuberculosis en Colombia. Biomédica 2004;24(Supl.):102-14, 2002.